鼎泽配资
鼎泽配资
肾单位是肾脏的基本 功能 单元,人类 双侧肾脏 共有 约 200 万个肾单位。 肾单位丢失是 导致 肾功能下降的主要原因【1】。由于负责产生哺乳动物肾单位的肾脏祖细胞在出生前后消失 , 发生肾损伤后人类肾脏只能修复损伤较小的肾单位,而不具备 再 生 全新 肾单位的能力 , 这也是肾脏 疾病 高发 且 难以 治愈的主要 原因【2】。 因此,学界一直希望寻找到可以在成体中再生肾单位的肾脏干细胞( Renal Stem Cells ,RSCs),为肾脏疾病的治疗提供新的方法。 然而, RSCs 的身份至今仍未被确证,是该领域长期未解的科学难题 。
202 5 年 8 月 2 2 日,陆军军医大学刘赤教授 、赵景宏教授团队在 S cience Advance s 期刊 在线发表了题为:
Identification of Renal Stem Cells in Zebrafish的研究论文。该研究 在斑马鱼中 , 通过单细胞 RNA 测序、光转换谱系追踪、干细胞连续移植等方法 ,鉴定到了一种由eya2six2apax2a等基因标记的RSCs类群,此类细胞通过分化增殖去分化模式进行更新,与经典干细胞的自我更新模式存在显著差异

与人类不同,成年斑马鱼具有强大的肾脏再生能力, 在急性肾损伤( Acute kidney injury , AKI )造成大量肾单位丢失后 ,斑马鱼能在 14 天内再生 一批全新 肾单位,恢复肾功能【4】。 既往的 研究表明, 斑马鱼成体肾脏中 存在 一种由 lhx1a 基因 标记的细胞团,通过移植该类细胞团可以再生新的肾单位【4, 5】。 但是,斑马鱼中是否存在 RSCs , RSCs 存在于 lhx1a + 细胞团中还是 为 产生 该 细胞团的前体细胞等问题并未有研究回答 。 RSCs 的产生、分化和更新方式 以及 调控 这些过程的 机制也未阐明。
因此,该研究 首先利用单细胞 RNA 测序的方法 , 对庆大霉素 肾 损伤后 四天的斑马鱼肾脏进行了分析。 发现 其中 一 个细胞群 (细胞群 15 ) 表达哺乳动物肾脏祖细胞的相关标记 基因 : six2a 、 pax2a 、 lhx1a 等。 使用 由 CRISPR/Cas9 基因敲入的方法构建的 Tg (six2a:eGFP) 转基因品系,可以在成体肾脏中发现两种类型的 eGFP + 细胞。一种为间充质状态的细胞,另一类为聚集成团的细胞,这些细胞团为 lhx1a 阳性 。通过进一 步 的亚群分析,可以发现细胞群 15 可以进一步分为两个亚群,亚群 一 高表达 six2a 、 eya2 以及间充质细胞标记 基因 。亚群 二 高表达 lhx1a 、 pax8 、 fgf8a 以及上皮细胞标记 基因 。 因此作者 推测亚群 一 为 RSCs ,而亚群 二 为 RSCs 分化而成的 细胞团组成细胞。
为了进一 步 确认 RSCs 的起源以及更新过程,作者尝试使用哺乳动物肾脏祖细胞 的早期 标记: osr1 、 lhx1a 、 eya1 进行追踪。但是,使用这三个基因的敲入 转基因 品系 均不能够标记 斑马鱼早期胚胎肾脏中 的 RSCs 。而后作者发现 RSCs 中 高表达 eya1 的同家族基因 eya2 。通过构建该基因的敲入 转基 因 品系 Tg ( eya2 :eGFP) , 发现在斑马鱼胚胎发育 46 小时( hours post fertilization , hpf ), eGFP + 细胞 在斑马鱼胚胎肾小管之间出现。这些细胞在 84 hpf 开始表达 Pax2a ,随后开始 six2a 的表达。这些细胞的体积 在 97 hpf 开始增大,并伸出伪足向肾小管上方迁移,移动到肾小管上方约 20 微米左右的位置。
继续追踪可以发现这些细胞在出生后 8 天 开始表达 lhx1a 。 与此前报道相同【5】, 它们 能通过聚集 形 成细胞团,并进行增殖和分化 产生 新的肾单位。作者进一步使用 EdU 细胞增殖检测,发现单个游离的 RSCs 并不具备增殖的能力,而聚集成团的细胞则具有较强的增殖 信号 。因此,作者推测 RSCs 的更新主要发生在细胞团中。为了追踪 RSCs 的更新过程,作者使用了光转换谱系追踪方法。通过将 Tg (six2a: Kaede ) 胚胎 中的单个细胞团进行变色和追踪, 可以发现单个细胞团可以分化为一个完整的肾单位,并且在肾单位的周围还存在由该细胞 团产生 的 4 - 5 个新生 RSCs 。 免疫荧光和原位杂交结果表明 , RSCs 在聚集成团后开始进行分化,细胞也通过间质上皮转化 ( Mesenchymal-Epithelial Transition , MET ) 形成细胞间的连接。随着细胞团的增殖,细胞团下方的细胞开始 高表达 lhx1a 并向肾单位分化 ,而细胞团顶部的细胞重新 开始 高表达 eya2 和 six2a ,随着进一步的发育,顶部的细胞开始脱离细胞团形成新的 RSCs 。这个过程说明 RSCs 的更新过程中发生了去分化以及上皮间质转化 ( Epithelial-Mesenchymal Transition , EMT ) 。
在未损伤的成体肾脏中,研究团队发现由约 12 个细胞组成的 eya2⁻ 细胞团长期处于静息状态。肾损伤发生后,部分细胞开始激活 eya2 ,继而表达 six2a 、 lhx1a 等基因,形成新的肾单位和 RSCs 。这一过程与胚胎期肾单位发育高度相似。连续移植实验进一步证明, six2a⁺/lhx1a⁻/cdh17⁻ 的 RSCs 具有长期产生肾单位并更新自身的能力。在 eya2 突变体中, RSCs 的更新显著减弱但未完全丧失;而 eya1 在其中发挥部分遗传代偿作用,但无法完全恢复更新功能,提示斑马鱼 RSCs 与 哺乳动物肾脏祖细胞在调控机制上存在差异【6】。 机制层面,免疫共沉淀实验显示 Eya2 通过与 Six1b 形成复合物发挥功能。既往研究表明 Six1 能通过激活 Pax2 和 Six2 的表达调控 哺乳动物 肾脏祖细胞发育【7】。同时, Wnt 信号在肾脏发育和再生中发挥关键作用【8】。研究发现, 过度激活或完全抑制 Wnt 信号均会阻断 RSCs 的更新。进一步实验证实, Wnt 信号的下游因子 Lef1 可直接结合 eya2 启动子并激活其转录,从而调控 RSCs 的更新 。
综上,该研究发现了斑马鱼肾脏中负责肾单位发育和再生的一类干细胞。这类干细胞的主要分子标记为eya2、pax2a、six2a。肾单位发育和再生过程中, 3-4 个 RSCs 通过聚集形成细胞团,并进行增殖和分化,细胞团的底部细胞分化为一个新的肾单位,而顶部细胞通过去分化产生新的 RSCs 。这种 “分化 - 增殖 - 去分化”的 更新 模式,显著区别于经典干细胞的自我更新 方式。
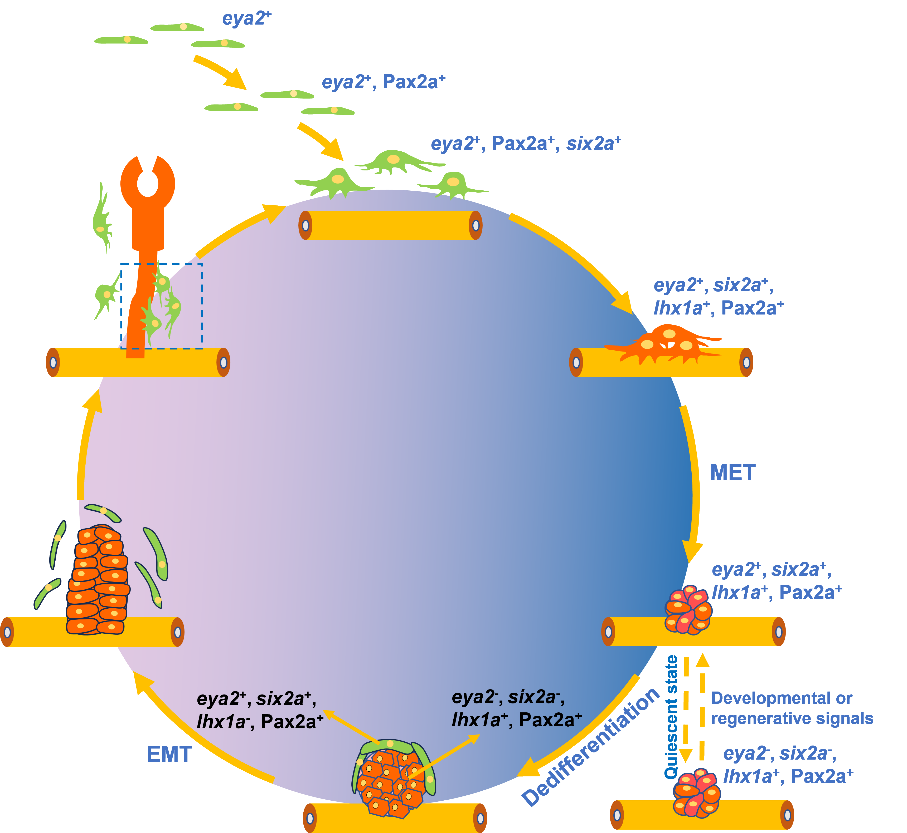
图 :肾脏干细胞更新机制图 。
该研究为探索RSCs产生和更新的机制提供了必要的前提条件,也为制造人类肾脏干细胞并治愈肾脏疾病带来了希望。
原文链接:http://doi.org/10.1126/sciadv.adx5296鼎泽配资
制版人: 十一
参考文献
1. P. Romagnani, G. Remuzzi, R. Glassock, A. Levin, K. J. Jager, M. Tonelli, Z. Massy, C. Wanner, H. J. Anders, Chronic kidney disease.
Nat Rev Dis Primers3, 17088 (2017).
2. Ronco C, Levin A, Warnock DG, Mehta R, Kellum JA, Shah S, et al. Improving outcomes from acute kidney injury (AKI): Report on an initiative.
Int J Artif Organs2007, 30(5): 373-376.
3. Yu T, Liu X, Tan X, Zhang Y, He Z, Yang W, Tian T, Li Y, Zhao J, Liu C. Identification of renal stem Cells in zebrafish.
Sci Adv2025, 10.1126/sciadv.adx5296.
4. Kamei CN, Liu Y, Drummond IA. Kidney Regeneration in Adult Zebrafish by Gentamicin Induced Injury.
J Vis Exp2015(102): e51912.
5. Diep CQ, Ma D, Deo RC, Holm TM, Naylor RW, Arora N, et al. Identification of adult nephron progenitors capable of kidney regeneration in zebrafish.
Nature2011鼎泽配资, 470(7332): 95-100.
6. J. Li, C. Cheng, J. Xu, T. Zhang, B. Tokat, G. Dolios, A. Ramakrishnan, L. Shen, R. Wang, P. X. Xu, The transcriptional coactivator Eya1 exerts transcriptional repressive activity by interacting with REST corepressors and REST-binding sequences to maintain nephron progenitor identity.
Nucleic Acids Res50, 10343-10359 (2022).
7 . P. X. Xu, W. Zheng, L. Huang, P. Maire, C. Laclef, D. Silvius, Six1 is required for the early organogenesis of mammalian kidney.
Development130, 3085-3094 (2003).
8. D. M. Iglesias, P. A. Hueber, L. Chu, R. Campbell, A. M. Patenaude, A. J. Dziarmaga, J. Quinlan, O. Mohamed, D. Dufort, P. R. Goodyer, Canonical WNT signaling during kidney development.
Am J Physiol Renal Physiol293, F494-500 (2007).
学术合作组织
(*排名不分先后)

战略合作伙伴
(*排名不分先后)
转载须知
【非原创文章】本文著作权归文章作者所有,欢迎个人转发分享,未经作者的允许禁止转载,作者拥有所有法定权利,违者必究。
BioArt
Med
Plants
人才招聘
近期直播推荐

点击主页推荐活动
关注更多最新活动!

在线配资提示:文章来自网络,不代表本站观点。